
OCTOBER 8, 2018. BY STAN GRANT.
Beginning in the late 1980’s, Merlot became a major fixture in the California wine grape industry largely because it lends itself to smooth, fruity, darkly colored, and approachable red wines.
Merlot is truly an international variety with significant plantings in Italy, Switzerland, Slovenia, Croatia, Macedonia, Moldova, Montenegro, Romania, Bulgaria, Greece, Turkey, Russia, Chile, Argentina, Australia, New Zealand, and Mexico, as well as in wine producing regions within the United States outside of California. France, though, is Merlot’s homeland and there it is officially called Merlot noir. It is widely planted in France, but is most closely associated with the Bordeaux. In that region, it is the base for the red wines of St. Emilion and Pomerol, as well as a common blending ingredient for Cabernet Sauvignon based wines.

Fig. 1: Mature Merlot clusters are naturally loose. (Progressive Viticulture©)

Fig. 2: Lateral shoots arise at most nodes of Merlot shoots. (Progressive Viticulture©). Click HERE to enlarge photo.
In California, Merlot grapevines typically grow with moderate high vigor, somewhat similar to Petite Sirah and Zinfandel. Several of Merlot’s viticultural traits, however, are distinctly different from other wine grape varieties common to California. Unlike the aforementioned Petite Sirah and Zinfandel, Merlot clusters are loose and low density (Fig. 1). Perhaps most unique among Merlot’s traits is the prolific growth of lateral shoots at nodal positions along the stems of main shoots (Fig. 2).
Presumably, the abundance of laterals is the result of weak apical dominance. Apical dominance is a physiological phenomenon in which the main shoot tip suppresses the buds, including lateral buds, below it through the production of hormones and their downward movement. Due to its comparatively thin, flexible main stems and the weight of lateral shoots, Merlot canopies tend to be drapey and dense, with shaded interiors (Fig. 3).

Fig. 3: Merlot shoots naturally drape downward, shading canopy interiors. (Progressive Viticulture©)
Based on these shoot growth characteristics, Merlot, more than any other widely planted variety, benefits from early season upward shoot positioning. Positioning the shoot apex in this way enhances hormone production in the main shoot tip and the downward movement of hormones that promote lateral bud dormancy. As a result, lateral shoot growth is inhibited and the need for leaf and lateral shoot removal in the fruit zone lessened.
However, even with upward shoot positioning, most Merlot vineyards benefit from fruit zone leaf and lateral shoot removal. These actions ensure adequate fruit zone exposure to light and lessen the risk of strong vegetative aromas in the harvested fruit.
Extensive lateral shoot growth has another effect – competition for internal vine resources. Each shoot tip (the main as well as the lateral shoot tips) competes with one another and as bloom approaches, with developing clusters. This competition contributes the low fruit set Merlot is famous for.
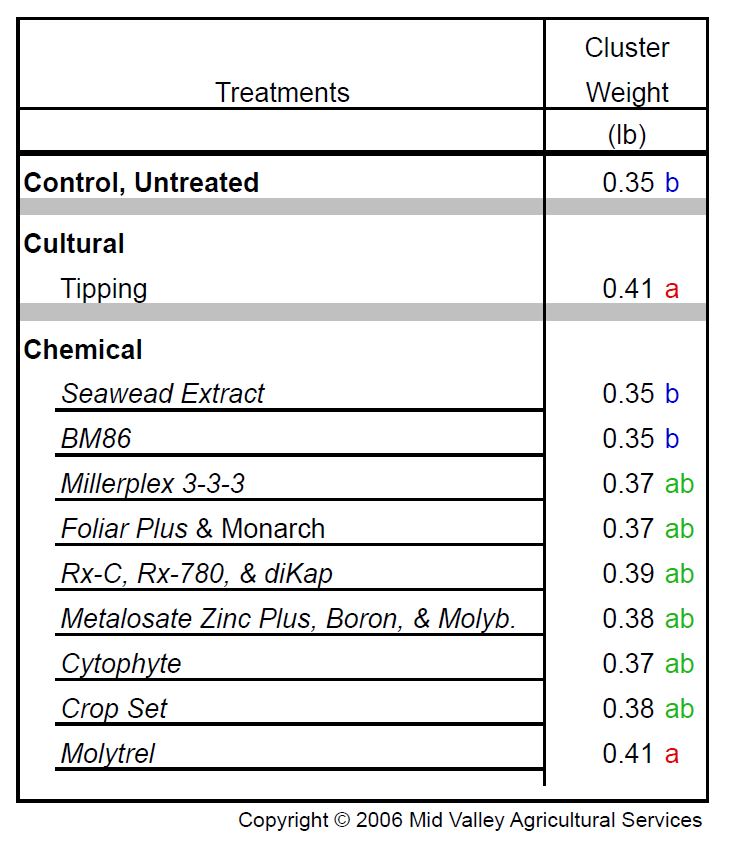
Table 1: Effect of shoot tipping and selected foliar applied products on Merlot cluster weight at harvest 2006. Treatments applied at first trace of bloom. Products applied at manufacturer’s recommended rates. (Source: Mid Valley Agricultural Services. Used with permission.) Click HERE to enlarge table.
Lateral shoot removal lessens the competition for internal resources and improves fruit set, but such a practice is too costly for most vineyards. However, Merlot growers have another option and that is a foliar application of molybdenum. Molybdenum is a plant micronutrient that is a cofactor for a number of biochemical processes in grapes, including the conversion of inorganic nitrogen taken up from the soil into organic forms in foliar tissues. In trials conducted by Mid Valley Agricultural Services, molybdenum was as effective as lateral removal for enhancing Merlot fruit set, especially if applied in quantities greater than 0.06 pounds per acre (Tables 1 and 2). Of course, this assumes vines are otherwise well nourished, especially with regard to carbohydrates and the mineral nutrients phosphorus, boron, and zinc.
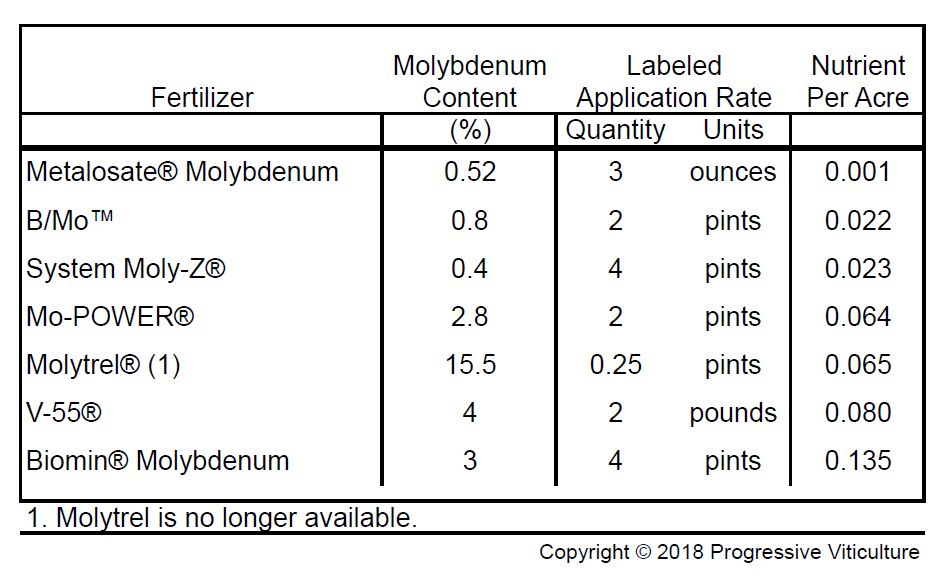
Table 2: Molybdenum content of selected fertilizers. (Progressive Viticulture©) Click HERE to enlarge table.
Internal competition may also be a factor in the poor stem lignification and associated high level of winterkill experienced in some Merlot vineyards. Encourage fully ripened, cold hardy stem wood through an integrated approach involving ample leaf exposure to sunlight, balanced crop and canopy growth, moderate water stress following fruit set, and a balanced supply of mineral nutrients, including adequate iron, manganese, copper, boron, and molybdenum.
Merlot appears more sensitive to water stress than many other red wine varieties. This apparent sensitivity is associated with the (anisohydric) behavior of tiny pores (stomata) on the undersides of leaves through which most vine water is lost. For Merlot, the guard cells at the pore openings are fairly insensitive to vine water stress. As such, Merlot leaves are prone to water stress induced leaf damage. To minimize such damage and ensure consistent production and timely fruit maturation, irrigate as needed to maintain moderate vine wine water stress (i.e. leaf water potential between -10 to -12 bars).
Thoughtful plant material selection can lessen the challenges of growing Merlot. There are several Merlot clones available. Among these, Merlot-08 and Merlot-10 tend to produce low yields due to fewer set berries per cluster. As a consequence, they are not suitable for many vineyards. Merlot-15 is the virus-tested selection of Merlot-181 from France, which is a consistent high yielding clone popular with many winemakers. Avoid rootstocks that tend to lessen fruit set, such as St. George and Dogridge, as well as those with low tolerance to drought, including 5C and 101-14 (for more information on rootstock selection, click HERE).
A version of this article was originally published in the Mid Valley Agricultural Services October 2012 newsletter and was updated for this blog post.
Further Reading
Benz, MJ; Anderson, MM; Williams, MA; Barnhisel, K; Wolpert, JA. Viticultural performance of five Merlot clones in Oakville, California. Am. J. Enol. Vitic. 57, 233-237. 2006.
Bettiga, LJ. Comparison of Merlot and Chardonnay clones in Monterey County: a preliminary report. In __ (Ed.). Int. Symp. Clonal Selection. pp. 93-95. 1995.
Brown, PH. Nutrient demand and fertilizer strategies: lessons learned from boron. In Conf. Proc. Calif. Plant Soil Conf., Modesto, Calif., Feb. 5-6, 2003. Calif. Chapt. Am. Soc. Agron. 2003.
Christensen, LP. Merlot. In Bettiga, LJ; Christensen, LP; Dokoozlian, NK; Golino, DA; McGourty, G; Smith, RJ; Verdegaal, PS; Walker, MA; Wolpert, JA; Weber, E. (Eds.). Wine Grape Varieties in California. University of California Agriculture and Natural Resources Publication 3419.
Fidelibus, MW; Christensen, LP; Katayama, DG; Verdenal, P-T; Cathline, K. Fruit characteristics of six Merlot grapevine selections in the Central San Joaquin Valley. Am. J. Enol. Vitic. 58, 259-261. 2007.
Guerra, B. Tipping Malbec shoots to improve set and yield. Practical Winery and Vineyard. Jan/Feb. 2006.
Kaiser, BN; Gridley, KL; Ngaire Brady, J; Phillips, T; Tyerman, SD. The role of molybdenum in agricultural plant production. Ann. Bot. 96, 745-754. 2005.
May, P. Flowering and fruit set in grapevines. Lythrum Press, Adelaide. 2004.
Pastena, B. Blind buds in the grapevine. 2. Blind buds and bio-ecological and agronomic factors. Vignevini. 17, 53-59. 1990.
Roberts, D; Blazer, R. The interaction of crop level and clone on the enologcial and viticultural characteristics of Merlot. In __ (Ed.). Int. Symp. Clonal Selection. pp. 133-136. 1995.
Robinson, J. Wines, grapes, and wines: the wine drinkers guide to grape varieties. Reed International Books, London, 1986.
Williams, C; Chvyl, L. Grapevine nutrition 8: molybdenum. Hartley Grove, South Australia, Cooperative Research Centre for Viticulture. 2006.
Williams, LE; Baeza, P. Relationships among ambient temperature and vapor pressure deficit and leaf and stem water potentials of fully irrigationed, field grown grapevines. Am. J. Enol. Vitic. 58, 173-181. 2007.
Have something interesting to say? Consider writing a guest blog article!
To subscribe to the Coffee Shop Blog, send an email to stephanie@lodiwine.com with the subject “blog subscribe.”
To join the Lodi Growers email list, send an email to stephanie@lodiwine.com with the subject “grower email subscribe” or click on “join our email list” to the right.
To receive Lodi Grower news and event promotions by mail, send your contact information to stephanie@lodiwine.com or call 209.367.4727.
For more information on the wines of Lodi, visit the Lodi Winegrape Commission’s consumer website, lodiwine.com.
